

Sestrin2 (SESN2): A Stress-Sensing Protein Linking Redox Control, mTOR Signaling, Ferroptosis, ER-Phagy, and Reproductive Protection
1. Introduction
Sestrin2 (SESN2) is a highly conserved stress-inducible protein that helps cells respond to oxidative, metabolic, hypoxic, and organelle-related injury (Lu et al., 2023; Ro et al., 2020). Because SESN2 responds to several forms of cellular injury, it is increasingly viewed as a regulator of stress adaptation in diseases involving oxidative damage, metabolic dysfunction, and chronic tissue injury (Lu et al., 2023; Rooban et al., 2024). In contrast to many proteins that work through a single specific signal transduction pathway, SESN2 works at a high level of cellular stress control, acting as a central hub to link many pathways, including antioxidant defenses, AMPK activation, mTOR complex 1 (mTORC1) inhibition, autophagy, mitophagy, and metabolic regulation (Ro et al., 2020). The broad range of signaling pathways associated with SESN2 helps explain its increasing number of studies on metabolic disorders, cardiovascular disease, liver disease, neurodegenerative disease, tissue injury, and cancer (Gong et al., 2021). What distinguishes SESN2 from other biomarkers is its unique ability to link the detection of cellular stress to several mechanisms involved in repairing damaged cells. While SESN2 indicates cellular stress, it also regulates how a cell responds to that stress (Lu et al., 2023). Therefore, unlike some single-function biomarkers, SESN2 can detect cellular stress and regulate how the cell adapts. Recent studies have associated SESN2 with ferroptosis, ER-phagy, and ovarian oxidative injury, further establishing it as an emerging protein in contemporary biomedical research (Zhang et al., 2024).
2. The Structure of SESN2
Human SESN2 consists of two main globin-like domains separated by an interdomain linker region. This structure allows it to bind with the proteins involved in redox control, amino acid sensing, and mTOR regulation. The N-terminal region has been linked to redox-related activity, whereas the C-terminal region contributes to leucine sensing and interaction with the GATOR2 complex (Lu et al., 2023; Valenstein et al., 2025). This organization is important for SESN2's role as a regulator of nutrient-sensitive growth signals. When leucine availability changes, SESN2-GATOR2 interactions help transmit amino acid sufficiency signals to mTORC1, thereby linking nutrient sensing with growth control (Haidurov and Budanov, 2024; Valenstein et al., 2025). This structural organization supports SESN2's role as a link between stress responses and nutrient-dependent growth regulation. Therefore, SESN2 should not be viewed as a passive indicator of stress. Its modular structure enables it to function as an interaction platform for other proteins. This platform connects oxidative stress, nutrient availability, autophagy, and mitochondrial quality control (Gong et al., 2021). This explains why dysfunction of SESN2 can contribute to many diseases rather than to a single isolated condition. As shown in Figure 1, the structural organization of SESN2 illustrates how a single stress-responsive protein can link redox-related residues, leucine sensing, and GATOR–mTOR signaling through distinct functional regions.
Figure 1. Structural organization and functional domains of human SESN2.
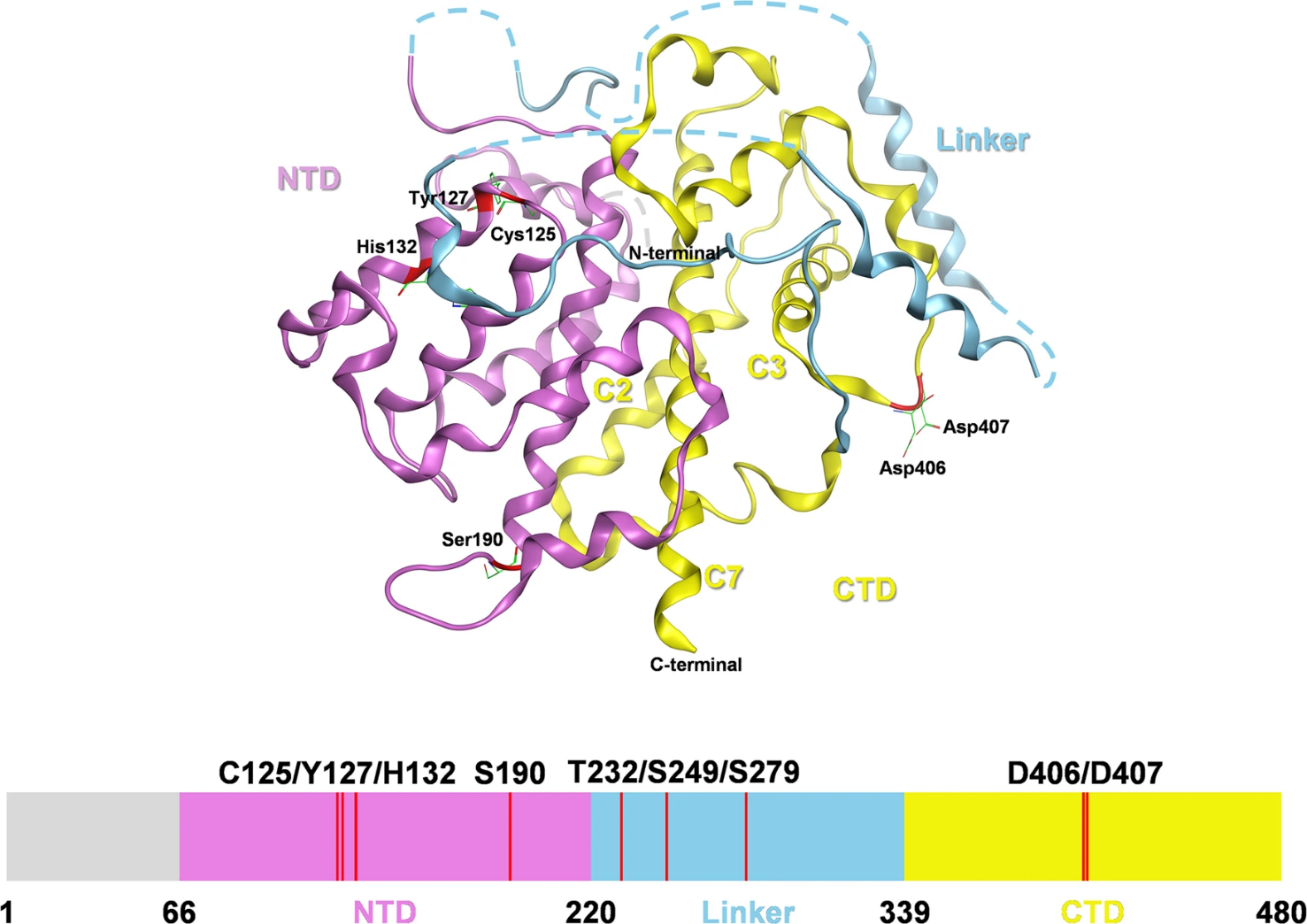
(Lu et al., 2023)
1. Regulation of SESN2 Expression
Several stress-responsive transcription factors regulate SESN2 expression. These include p53, hypoxia-inducible factor-1, nuclear factor erythroid 2-related factor 2, activating transcription factor 4, and activating transcription factor 6 (Lu et al., 2023). This broad regulation enables activation in response to various types of injury, including oxidative stress, hypoxia, DNA damage, and endoplasmic reticulum stress. One of the most important pathways that regulates SESN2 in oxidative injury is the NRF2 pathway. This pathway regulates antioxidant gene expression, while SESN2 has been shown to enhance NRF2-related defense mechanisms through p62-Keap1 interactions (Gong et al., 2021). This feedback mechanism links SESN2 induction to NRF2-related antioxidant signaling, enabling cells to enhance redox defense under oxidative stress (Gong et al., 2021; Li et al., 2025). SESN2 is closely associated with the AMPK-mTORC1 pathway. When a cell experiences stress, SESN2 activates AMPK and inhibits mTORC1 activity, thereby reducing unnecessary cellular proliferation and supporting cellular repair (Ro et al., 2020). This is biologically important because cells under stress should not continue to grow uncontrollably. Instead, they need to conserve energy, remove damaged components, and restore homeostasis.
2. SESN2 as a Redox and Metabolic Control Hub
One of the main biological roles of SESN2 is to regulate redox homeostasis. Redox imbalance (or oxidative stress) occurs when reactive oxygen species (ROS) are produced in excess of what an organism's antioxidant systems can neutralize. Excess ROS can damage DNA, proteins, lipids, and organelles, while NRF2-mediated antioxidant responses help maintain cellular redox homeostasis (Li et al., 2025). SESN2 plays a role in reducing oxidative injury by activating protective signaling networks. It supports antioxidant responses, reduces ROS accumulation, and promotes the removal of damaged mitochondria through mitophagy (Gong et al., 2021). As illustrated in Figure 2, SESN2 regulates cellular adaptation by linking oxidative stress responses to NRF2 activation, AMPK signaling, mTORC1 inhibition, and autophagy regulation. This is important because damaged mitochondria can be significant producers of ROS, worsening cellular injury. Beyond redox regulation, SESN2 also contributes to metabolic homeostasis by controlling mTORC1 activity. Under conditions of nutrient abundance, mTORC1 activity drives protein synthesis and cell growth. However, under stress conditions, SESN2 helps inhibit mTORC1 activity, thereby promoting autophagy for stress adaptation (Ro et al., 2020). Therefore, SESN2 is positioned as the regulatory component that balances the processes of cell growth and protection.
Figure 2. SESN2 as a stress-signaling hub linking NRF2, AMPK–mTOR and autophagy.
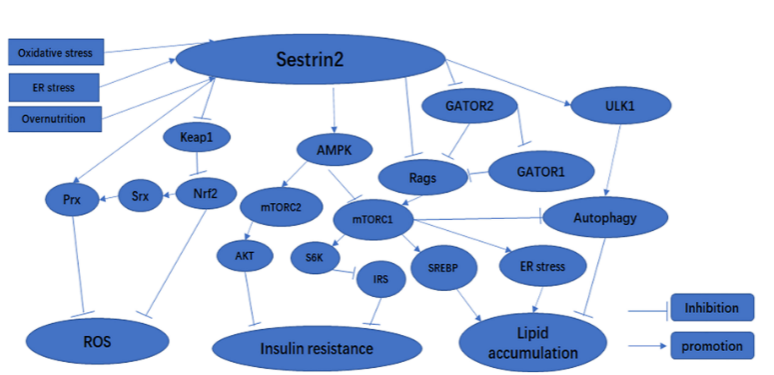
(Gong et al., 2021)
1. SESN2 in Autophagy, Mitophagy, and Cellular Quality Control
Autophagy is a protective process that removes damaged proteins and organelles. SESN2 supports autophagy by helping suppress mTORC1 and activate pathways that promote cellular recycling (Ro et al., 2020). The role of SESN2 in autophagy becomes even more critical with increasing exposure to oxidative stress, nutrient starvation, or other forms of metabolic injury. Mitophagy is a selective autophagy pathway that removes damaged or dysfunctional mitochondria to preserve mitochondrial quality control (M. Yang et al., 2024). SESN2 has been shown to play roles in both PINK1/Parkin-mediated mitochondrial quality control and the elimination of mitochondria that produce ROS (Lu et al., 2023). This process is important because damaged mitochondria can amplify ROS production, inflammatory signaling, and regulated cell-death pathways (M. Yang et al., 2024). Thus, SESN2 can support cellular protection under moderate stress by coordinating autophagy and mitophagy. However, its impact depends heavily on both the affected tissue(s) and the nature of the disease. Thus, depending on the conditions present in a specific disease state, SESN2 can protect cells from disease and maintain the survival of damaged or malignant cells (Gong et al., 2021). Therefore, SESN2 should be viewed as a context-specific regulatory element rather than a universal protective agent.
2. SESN2 and Ferroptosis Regulation
Ferroptosis is a regulated form of cell death driven by excessive lipid peroxidation and dependent on iron. Ferroptosis contributes to several pathological conditions, including ischemia-reperfusion injury, neurodegeneration, metabolic disease, and cancer, mainly through iron-dependent lipid peroxidation and failure of antioxidant defense (Chen et al., 2024). While SESN2 does not function as a classical ferroptosis enzyme like GPX4, it may modulate ferroptosis through antioxidant and stress-response mechanisms. Experimental studies have demonstrated that SESN2 can reduce ferroptosis through the Keap1-Nrf2 pathway. During intestinal ischemia-reperfusion injury, activation of SESN2 reduced ferroptosis-related damage and increased cellular protective responses (Zhang et al., 2024). This suggests that SESN2 may protect tissues by improving antioxidant potential and reducing lipid peroxidation. SESN2 has also been studied within diabetic retinopathy. Within this model, overexpression promoted cell viability, reduced apoptosis, inhibited ferroptosis, and stimulated autophagy (Xi et al., 2024). Because autophagy can either suppress or promote ferroptosis depending on cell type and disease context, SESN2-related ferroptosis regulation should be interpreted as context-dependent rather than universally protective (Liu et al., 2025).
3. SESN2 in ER Stress and ER-Phagy
The endoplasmic reticulum is primarily involved in protein folding, lipid processing, and the maintenance of cellular calcium levels. Cells activate an unfolded protein response when high levels of misfolded proteins accumulate in the ER. The goal of this response is to assist the cell’s recovery from ER-associated dysfunction (Lu et al., 2023). Failure to recover from ER stress may activate mechanisms that lead to apoptosis, inflammation, and tissue injury. Recent studies have demonstrated that SESN2 is also involved in ER-phagy. ER-phagy is an autophagic process that selectively removes damaged regions of the endoplasmic reticulum. ER-phagy contributes to ER proteostasis by targeting damaged ER fragments, misfolded proteins and ER-associated aggregates for lysosomal degradation (Wu et al., 2025). It has also been demonstrated that SESN2 may support ER-phagy through a biochemical pathway involving XBP1, mTORC1, TFEB/TFE3, and FAM134B. Therefore, this further defines the biological role(s) of SESN2 by indicating that it can also contribute to cellular protection against endoplasmic reticulum dysfunction during excessive misfolding (De Leonibus et al., 2024). During protein-misfolding stress, SESN2 induction can inhibit mTORC1-dependent phosphorylation of TFEB/TFE3, allowing these transcription factors to promote FAM134B-mediated ER-phagy (Figure 3).
Figure 3. SESN2-driven ER-phagy during protein-misfolding stress.
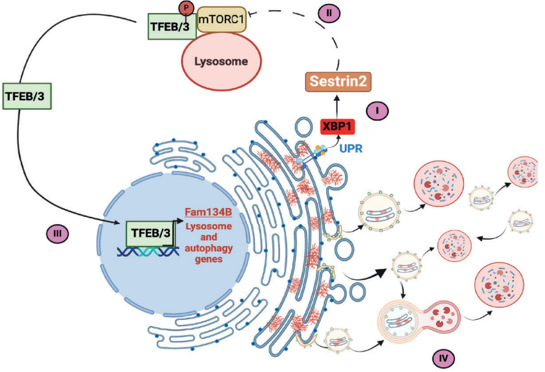
(De Leonibus et al., 2024)
1. Clinical Significance of SESN2 as a Disease Biomarker
SESN2 is gaining interest as a measurable disease-related protein. Because SESN2 is responsive to oxidative stress, inflammation, and tissue injury, changes in its levels could indicate cellular stress. It has been examined as a candidate biomarker in neurological injury and cardiometabolic disease, although its interpretation appears to depend on disease stage, tissue, and clinical context (Abdelsalam et al., 2024; Dou et al., 2023; Rooban et al., 2024). Serum levels of SESN2 were correlated with the severity of disease and functional outcomes following acute intracerebral hemorrhage. Higher serum SESN2 was associated with greater neurological deterioration and poorer prognosis (Dou et al., 2023). This suggests that circulating SESN2 levels may reflect the extent of tissue injury resulting from acute cerebral damage. However, SESN2 should not be considered a singular diagnostic indicator. Its expression can vary in response to numerous disease states, including hypoxia, oxidative stress, inflammation, and metabolic imbalances (Lu et al., 2023). Therefore, circulating SESN2 is best interpreted as a supportive stress-related marker rather than a stand-alone diagnostic test (Abdelsalam et al., 2024; Dou et al., 2023).
2. SESN2 in Ovarian Oxidative Stress and Reproductive Protection
The reproductive relevance of SESN2 is emerging, particularly regarding ovarian oxidative stress, mitochondrial function, and granulosa cell survival (Huang et al., 2024; Wang et al., 2023). Ovarian function depends strongly on mitochondrial activity, redox balance, oocyte quality, and granulosa-cell survival (Ju et al., 2024; Z. Yang et al., 2024). When oxidative stress increases, granulosa cells may undergo apoptosis, follicular development may be disrupted, and female fertility may decline. Some studies found high levels of miR-484 in both ovarian and granulosa cell models subjected to oxidative stress. Furthermore, the LINC00958/miR-484/SESN2 axis was also found to regulate mitochondrial function and mitochondrial-related apoptosis in granulosa cells (Wang et al., 2023). These findings suggest that SESN2 may protect granulosa cells under oxidative stress, but this should be interpreted cautiously because whole animal fertility outcomes may not always change after Sestrin gene loss (Wang et al., 2023; Wang et al., 2024). Other studies have shown that proanthocyanidin compounds protect against the 3-NPA-induced decline in ovarian function by activating the SESTRIN2–NRF2 pathway. In ovarian oxidative injury, activation of the SESTRIN2–NRF2 pathway reduces ROS accumulation, supports mitochondrial protection, and helps preserve ovarian function (Huang et al., 2024). These findings link SESN2 to natural antioxidant-mediated ovarian protection.
Figure 4. Sestrin2–NRF2 signaling in ovarian oxidative-stress protection.
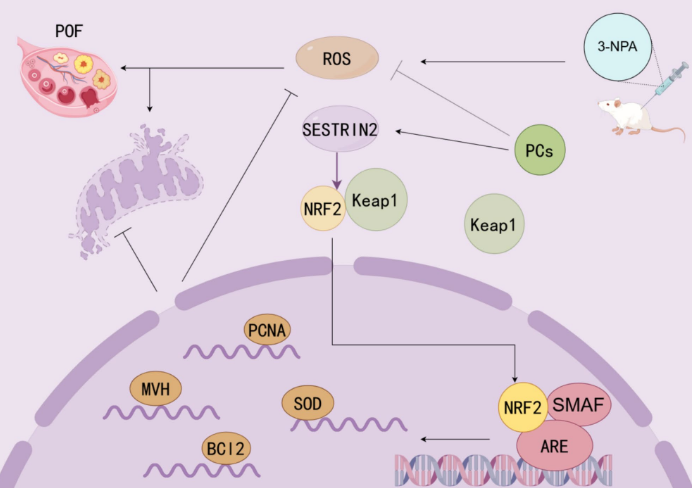
(Huang et al., 2024)
1. Methods for Detecting and Analyzing SESN2
SESN2 can be evaluated at the mRNA, protein, and functional levels. RT-qPCR is most commonly used to evaluate SESN2 gene expression, while Western blotting assesses SESN2 protein abundance (Huang et al., 2024). Immunohistochemistry and immunofluorescence are typically used to localize SESN2 to specific tissues or cellular compartments. Functional studies often require SESN2 knockdown or overexpression. These approaches help determine whether SESN2 is only associated with disease or whether it directly contributes to protection or injury (Xi et al., 2024). Mechanistic studies commonly combine SESN2 knockdown or overexpression with ROS assays, apoptosis analysis, mitochondrial membrane-potential detection, autophagy markers, and ferroptosis-related readouts. In addition to assessing antioxidant activity, endpoints for reproductive research studies involving SESN2 may include granulosa cell viability, mitochondrial function, apoptosis, ovarian histology, hormone levels, and oxidative stress biomarkers (Wang et al., 2023). These endpoints can help assess whether SESN2 improves ovarian-cell survival and protects against ovarian functional decline. In natural-compound studies, SESN2 can also be tested as a pathway target for antioxidants such as proanthocyanidins (Huang et al., 2024).
2. Therapeutic Potential and Future Perspectives
SESN2’s therapeutic effects arise from its position upstream of multiple protective mechanisms. Activation of SESN2 may mitigate oxidative stress, ferroptosis, inflammation, mitochondrial damage, and endoplasmic reticulum (ER) stress (Lu et al., 2023). Thus, SESN2 is better described as a potential therapeutic target that still requires disease-specific validation (Rooban et al., 2024; Wang et al., 2024). In addition to metabolic disease, SESN2 may be involved in regulating insulin sensitivity, lipid metabolism, and energy balance. Its ability to influence AMPK, mTORC1, and autophagy provides relevance to diabetes, obesity, fatty liver disease, and cardiac injury (Gong et al., 2021). However, because SESN2 has varying effects depending on the specific tissue or disease condition, careful consideration is required when designing therapeutic approaches. In reproductive biology, SESN2 should be investigated further using models that connect granulosa-cell survival, oocyte competence, ovarian reserve, and fertility outcomes under oxidative stress (Wang et al., 2024; Wang et al., 2023; Yang et al., 2024). It would also be valuable to test whether natural compounds can protect ovarian function through SESN2–NRF2, SESN2–AMPK, or SESN2–mTOR pathways (Huang et al., 2024).
3. Conclusion
SESN2 is an important stress-responsive protein that helps maintain cellular homeostasis during oxidative, metabolic, and organelle-related injury. It supports antioxidant defense and regulates AMPK-mTOR signaling, autophagy, mitophagy, ferroptosis, and endoplasmic reticulum (ER) quality control. Through these mechanisms, SESN2 can reduce ROS production, limit lipid peroxidation, promote the removal of damaged organelles, and improve cellular adaptation to stress. Its reproductive relevance is also important because ovarian function depends on mitochondrial activity, granulosa cell survival, and redox stability. SESN2-regulated pathways may therefore contribute to ovarian protection under oxidative stress, although direct effects on fertility outcomes still require further confirmation.
References
Abdelsalam, S.S., Zahid, M.A., Raïq, H., Abunada, H., Elsayed, A.E., Parray, A. and Agouni, A. (2024). The association between plasma levels of Sestrin2 and risk factors of cardiovascular diseases in healthy and diabetic adults: a study of Qatar Biobank data. Biomolecules and Biomedicine, 25(7), pp.1479–1490. https://doi.org/10.17305/bb.2024.11418.
Chen, F., Kang, R., Tang, D. and Liu, J. (2024). Ferroptosis: principles and significance in health and disease. Journal of Hematology & Oncology, 17, 41. https://doi.org/10.1186/s13045-024-01564-3.
De Leonibus, C., Maddaluno, M., Ferriero, R., Besio, R., Cinque, L., Lim, P.J., Palma, A., De Cegli, R., Gagliotta, S., Montefusco, S., Iavazzo, M., Rohrbach, M., Giunta, C., Polishchuk, E., Medina, D.L., Di Bernardo, D., Forlino, A., Piccolo, P. and Settembre, C. (2024). Sestrin2 drives ER-phagy in response to protein misfolding. Developmental Cell, 59, pp.2035–2052.e10. https://doi.org/10.1016/j.devcel.2024.07.004.
Dou, X., Dong, W., Gu, Y., Zhang, T. and Zhang, J. (2023). Significance of serum sestrin2 as a biomarker of severity and functional outcome in acute intracerebral hemorrhage: a prospective observational longitudinal study. BMC Neurology, 23, 424. https://doi.org/10.1186/s12883-023-03470-6.
Gong, L., Wang, Z.Z., Wang, Z.G. and Zhang, Z. (2021). Sestrin2 as a potential target for regulating metabolic-related diseases. Frontiers in Endocrinology, 12, 751020. https://doi.org/10.3389/fendo.2021.751020.
Haidurov, A. and Budanov, A.V. (2024). Locked in structure: Sestrin and GATOR—a billion-year marriage. Cells, 13, 1587. https://doi.org/10.3390/cells13181587.
Huang, Y., Cui, Y., Huang, J., Huang, X., Wang, Z., Luo, T. and Li, J. (2024). Proanthocyanidins protects 3-NPA-induced ovarian function decline by activating SESTRIN2-NRF2-mediated oxidative stress in mice. Scientific Reports, 14, 25643. https://doi.org/10.1038/s41598-024-76743-w.
Ju, W., Zhao, Y., Yu, Y., Zhao, S. and Lian, F. (2024). Mechanisms of mitochondrial dysfunction in ovarian aging and potential interventions. Frontiers in Endocrinology, 15, 1361289. https://doi.org/10.3389/fendo.2024.1361289.
Li, B., Ming, H., Qin, S., Nice, E.C., Dong, J., Du, Z. and Huang, C. (2025). Redox regulation: mechanisms, biology and therapeutic targets in diseases. Signal Transduction and Targeted Therapy, 10, 72. https://doi.org/10.1038/s41392-024-02095-6.
Liu, X., Tuerxun, H., Zhao, Y.X., Li, Y., Wen, S., Li, X. and Zhao, Y.G. (2025). Crosstalk between ferroptosis and autophagy: broaden horizons of cancer therapy. Journal of Translational Medicine, 23, 18. https://doi.org/10.1186/s12967-024-06059-w.
Lu, C., Jiang, Y., Xu, W. and Bao, X. (2023). Sestrin2: multifaceted functions, molecular basis, and its implications in liver diseases. Cell Death & Disease, 14, 160. https://doi.org/10.1038/s41419-023-05669-4.
Ro, S.-H., Fay, J., Cyuzuzo, C.I., Jang, Y., Lee, N., Song, H.-S. and Harris, E.N. (2020). SESTRINs: emerging dynamic stress-sensors in metabolic and environmental health. Frontiers in Cell and Developmental Biology, 8, 603421. https://doi.org/10.3389/fcell.2020.603421.
Rooban, S., Senghor, K.A.A., Vinodhini, V.M. and Kumar, J.S. (2024). Sestrin2 at the crossroads of cardiovascular disease and diabetes: a comprehensive review. Obesity Medicine, 51, 100558. https://doi.org/10.1016/j.obmed.2024.100558.
Valenstein, M.L., Wranik, M., Lalgudi, P.V., Linde-Garelli, K.Y., Choi, Y., Chivukula, R.R., Sabatini, D.M. and Rogala, K.B. (2025). Structural basis for the dynamic regulation of mTORC1 by amino acids. Nature, 646, pp.493–500. https://doi.org/10.1038/s41586-025-09428-7.
Wang, M., Chen, W., Zeng, X., Wang, T., Sun, Y. and Yang, Q. (2024). Sestrin1, 2, and 3 are dispensable for female fertility in mice. Journal of Ovarian Research, 17, 28. https://doi.org/10.1186/s13048-024-01345-z.
Wang, X., Yang, J., Li, H.Y., Mu, H., Zeng, L., Cai, S., Su, P., Li, H.B., Zhang, L. and Xiang, W. (2023). miR-484 mediates oxidative stress-induced ovarian dysfunction and promotes granulosa cell apoptosis via SESN2 downregulation. Redox Biology, 62, 102684. https://doi.org/10.1016/j.redox.2023.102684.
Wu, S.A., Li, Z.J. and Qi, L. (2025). Endoplasmic reticulum (ER) protein degradation by ER-associated degradation and ER-phagy. Trends in Cell Biology, 35, pp.576–591. https://doi.org/10.1016/j.tcb.2025.01.002.
Xi, X., Chen, Q., Ma, J., Wang, X., Zhang, J. and Li, Y. (2024). Sestrin2 ameliorates diabetic retinopathy by regulating autophagy and ferroptosis. Journal of Molecular Histology, 55, pp.169–184. https://doi.org/10.1007/s10735-023-10180-3.
Yang, M., Wei, X., Yi, X. and Jiang, D.-S. (2024). Mitophagy-related regulated cell death: molecular mechanisms and disease implications. Cell Death & Disease, 15, 505. https://doi.org/10.1038/s41419-024-06804-5.
Yang, Z., Liu, S. and Pan, X. (2024). Research progress on mitochondrial damage and repairing in oocytes: a review. Mitochondrion, 75, 101845. https://doi.org/10.1016/j.mito.2024.101845.
Zhang, L., Ding, K., Liao, S., Zhang, Y., Liao, H., Chen, R. and Meng, Q. (2024). Sestrin2 reduces ferroptosis via the Keap1/Nrf2 signaling pathway after intestinal ischemia-reperfusion. Free Radical Biology and Medicine, 214, pp.115–128. https://doi.org/10.1016/j.freeradbiomed.2024.02.003.
Sestrin2 (SESN2) is a highly conserved stress-inducible protein that helps cells respond to oxidative, metabolic, hypoxic, and organelle-related injury (Lu et al., 2023; Ro et al., 2020). Because SESN2 responds to several forms of cellular injury, it is increasingly viewed as a regulator of stress adaptation in diseases involving oxidative damage, metabolic dysfunction, and chronic tissue injury (Lu et al., 2023; Rooban et al., 2024). In contrast to many proteins that work through a single specific signal transduction pathway, SESN2 works at a high level of cellular stress control, acting as a central hub to link many pathways, including antioxidant defenses, AMPK activation, mTOR complex 1 (mTORC1) inhibition, autophagy, mitophagy, and metabolic regulation (Ro et al., 2020). The broad range of signaling pathways associated with SESN2 helps explain its increasing number of studies on metabolic disorders, cardiovascular disease, liver disease, neurodegenerative disease, tissue injury, and cancer (Gong et al., 2021). What distinguishes SESN2 from other biomarkers is its unique ability to link the detection of cellular stress to several mechanisms involved in repairing damaged cells. While SESN2 indicates cellular stress, it also regulates how a cell responds to that stress (Lu et al., 2023). Therefore, unlike some single-function biomarkers, SESN2 can detect cellular stress and regulate how the cell adapts. Recent studies have associated SESN2 with ferroptosis, ER-phagy, and ovarian oxidative injury, further establishing it as an emerging protein in contemporary biomedical research (Zhang et al., 2024).
2. The Structure of SESN2
Human SESN2 consists of two main globin-like domains separated by an interdomain linker region. This structure allows it to bind with the proteins involved in redox control, amino acid sensing, and mTOR regulation. The N-terminal region has been linked to redox-related activity, whereas the C-terminal region contributes to leucine sensing and interaction with the GATOR2 complex (Lu et al., 2023; Valenstein et al., 2025). This organization is important for SESN2's role as a regulator of nutrient-sensitive growth signals. When leucine availability changes, SESN2-GATOR2 interactions help transmit amino acid sufficiency signals to mTORC1, thereby linking nutrient sensing with growth control (Haidurov and Budanov, 2024; Valenstein et al., 2025). This structural organization supports SESN2's role as a link between stress responses and nutrient-dependent growth regulation. Therefore, SESN2 should not be viewed as a passive indicator of stress. Its modular structure enables it to function as an interaction platform for other proteins. This platform connects oxidative stress, nutrient availability, autophagy, and mitochondrial quality control (Gong et al., 2021). This explains why dysfunction of SESN2 can contribute to many diseases rather than to a single isolated condition. As shown in Figure 1, the structural organization of SESN2 illustrates how a single stress-responsive protein can link redox-related residues, leucine sensing, and GATOR–mTOR signaling through distinct functional regions.
Figure 1. Structural organization and functional domains of human SESN2.
(Lu et al., 2023)
1. Regulation of SESN2 Expression
Several stress-responsive transcription factors regulate SESN2 expression. These include p53, hypoxia-inducible factor-1, nuclear factor erythroid 2-related factor 2, activating transcription factor 4, and activating transcription factor 6 (Lu et al., 2023). This broad regulation enables activation in response to various types of injury, including oxidative stress, hypoxia, DNA damage, and endoplasmic reticulum stress. One of the most important pathways that regulates SESN2 in oxidative injury is the NRF2 pathway. This pathway regulates antioxidant gene expression, while SESN2 has been shown to enhance NRF2-related defense mechanisms through p62-Keap1 interactions (Gong et al., 2021). This feedback mechanism links SESN2 induction to NRF2-related antioxidant signaling, enabling cells to enhance redox defense under oxidative stress (Gong et al., 2021; Li et al., 2025). SESN2 is closely associated with the AMPK-mTORC1 pathway. When a cell experiences stress, SESN2 activates AMPK and inhibits mTORC1 activity, thereby reducing unnecessary cellular proliferation and supporting cellular repair (Ro et al., 2020). This is biologically important because cells under stress should not continue to grow uncontrollably. Instead, they need to conserve energy, remove damaged components, and restore homeostasis.
2. SESN2 as a Redox and Metabolic Control Hub
One of the main biological roles of SESN2 is to regulate redox homeostasis. Redox imbalance (or oxidative stress) occurs when reactive oxygen species (ROS) are produced in excess of what an organism's antioxidant systems can neutralize. Excess ROS can damage DNA, proteins, lipids, and organelles, while NRF2-mediated antioxidant responses help maintain cellular redox homeostasis (Li et al., 2025). SESN2 plays a role in reducing oxidative injury by activating protective signaling networks. It supports antioxidant responses, reduces ROS accumulation, and promotes the removal of damaged mitochondria through mitophagy (Gong et al., 2021). As illustrated in Figure 2, SESN2 regulates cellular adaptation by linking oxidative stress responses to NRF2 activation, AMPK signaling, mTORC1 inhibition, and autophagy regulation. This is important because damaged mitochondria can be significant producers of ROS, worsening cellular injury. Beyond redox regulation, SESN2 also contributes to metabolic homeostasis by controlling mTORC1 activity. Under conditions of nutrient abundance, mTORC1 activity drives protein synthesis and cell growth. However, under stress conditions, SESN2 helps inhibit mTORC1 activity, thereby promoting autophagy for stress adaptation (Ro et al., 2020). Therefore, SESN2 is positioned as the regulatory component that balances the processes of cell growth and protection.
Figure 2. SESN2 as a stress-signaling hub linking NRF2, AMPK–mTOR and autophagy.
(Gong et al., 2021)
1. SESN2 in Autophagy, Mitophagy, and Cellular Quality Control
Autophagy is a protective process that removes damaged proteins and organelles. SESN2 supports autophagy by helping suppress mTORC1 and activate pathways that promote cellular recycling (Ro et al., 2020). The role of SESN2 in autophagy becomes even more critical with increasing exposure to oxidative stress, nutrient starvation, or other forms of metabolic injury. Mitophagy is a selective autophagy pathway that removes damaged or dysfunctional mitochondria to preserve mitochondrial quality control (M. Yang et al., 2024). SESN2 has been shown to play roles in both PINK1/Parkin-mediated mitochondrial quality control and the elimination of mitochondria that produce ROS (Lu et al., 2023). This process is important because damaged mitochondria can amplify ROS production, inflammatory signaling, and regulated cell-death pathways (M. Yang et al., 2024). Thus, SESN2 can support cellular protection under moderate stress by coordinating autophagy and mitophagy. However, its impact depends heavily on both the affected tissue(s) and the nature of the disease. Thus, depending on the conditions present in a specific disease state, SESN2 can protect cells from disease and maintain the survival of damaged or malignant cells (Gong et al., 2021). Therefore, SESN2 should be viewed as a context-specific regulatory element rather than a universal protective agent.
2. SESN2 and Ferroptosis Regulation
Ferroptosis is a regulated form of cell death driven by excessive lipid peroxidation and dependent on iron. Ferroptosis contributes to several pathological conditions, including ischemia-reperfusion injury, neurodegeneration, metabolic disease, and cancer, mainly through iron-dependent lipid peroxidation and failure of antioxidant defense (Chen et al., 2024). While SESN2 does not function as a classical ferroptosis enzyme like GPX4, it may modulate ferroptosis through antioxidant and stress-response mechanisms. Experimental studies have demonstrated that SESN2 can reduce ferroptosis through the Keap1-Nrf2 pathway. During intestinal ischemia-reperfusion injury, activation of SESN2 reduced ferroptosis-related damage and increased cellular protective responses (Zhang et al., 2024). This suggests that SESN2 may protect tissues by improving antioxidant potential and reducing lipid peroxidation. SESN2 has also been studied within diabetic retinopathy. Within this model, overexpression promoted cell viability, reduced apoptosis, inhibited ferroptosis, and stimulated autophagy (Xi et al., 2024). Because autophagy can either suppress or promote ferroptosis depending on cell type and disease context, SESN2-related ferroptosis regulation should be interpreted as context-dependent rather than universally protective (Liu et al., 2025).
3. SESN2 in ER Stress and ER-Phagy
The endoplasmic reticulum is primarily involved in protein folding, lipid processing, and the maintenance of cellular calcium levels. Cells activate an unfolded protein response when high levels of misfolded proteins accumulate in the ER. The goal of this response is to assist the cell’s recovery from ER-associated dysfunction (Lu et al., 2023). Failure to recover from ER stress may activate mechanisms that lead to apoptosis, inflammation, and tissue injury. Recent studies have demonstrated that SESN2 is also involved in ER-phagy. ER-phagy is an autophagic process that selectively removes damaged regions of the endoplasmic reticulum. ER-phagy contributes to ER proteostasis by targeting damaged ER fragments, misfolded proteins and ER-associated aggregates for lysosomal degradation (Wu et al., 2025). It has also been demonstrated that SESN2 may support ER-phagy through a biochemical pathway involving XBP1, mTORC1, TFEB/TFE3, and FAM134B. Therefore, this further defines the biological role(s) of SESN2 by indicating that it can also contribute to cellular protection against endoplasmic reticulum dysfunction during excessive misfolding (De Leonibus et al., 2024). During protein-misfolding stress, SESN2 induction can inhibit mTORC1-dependent phosphorylation of TFEB/TFE3, allowing these transcription factors to promote FAM134B-mediated ER-phagy (Figure 3).
Figure 3. SESN2-driven ER-phagy during protein-misfolding stress.
(De Leonibus et al., 2024)
1. Clinical Significance of SESN2 as a Disease Biomarker
SESN2 is gaining interest as a measurable disease-related protein. Because SESN2 is responsive to oxidative stress, inflammation, and tissue injury, changes in its levels could indicate cellular stress. It has been examined as a candidate biomarker in neurological injury and cardiometabolic disease, although its interpretation appears to depend on disease stage, tissue, and clinical context (Abdelsalam et al., 2024; Dou et al., 2023; Rooban et al., 2024). Serum levels of SESN2 were correlated with the severity of disease and functional outcomes following acute intracerebral hemorrhage. Higher serum SESN2 was associated with greater neurological deterioration and poorer prognosis (Dou et al., 2023). This suggests that circulating SESN2 levels may reflect the extent of tissue injury resulting from acute cerebral damage. However, SESN2 should not be considered a singular diagnostic indicator. Its expression can vary in response to numerous disease states, including hypoxia, oxidative stress, inflammation, and metabolic imbalances (Lu et al., 2023). Therefore, circulating SESN2 is best interpreted as a supportive stress-related marker rather than a stand-alone diagnostic test (Abdelsalam et al., 2024; Dou et al., 2023).
2. SESN2 in Ovarian Oxidative Stress and Reproductive Protection
The reproductive relevance of SESN2 is emerging, particularly regarding ovarian oxidative stress, mitochondrial function, and granulosa cell survival (Huang et al., 2024; Wang et al., 2023). Ovarian function depends strongly on mitochondrial activity, redox balance, oocyte quality, and granulosa-cell survival (Ju et al., 2024; Z. Yang et al., 2024). When oxidative stress increases, granulosa cells may undergo apoptosis, follicular development may be disrupted, and female fertility may decline. Some studies found high levels of miR-484 in both ovarian and granulosa cell models subjected to oxidative stress. Furthermore, the LINC00958/miR-484/SESN2 axis was also found to regulate mitochondrial function and mitochondrial-related apoptosis in granulosa cells (Wang et al., 2023). These findings suggest that SESN2 may protect granulosa cells under oxidative stress, but this should be interpreted cautiously because whole animal fertility outcomes may not always change after Sestrin gene loss (Wang et al., 2023; Wang et al., 2024). Other studies have shown that proanthocyanidin compounds protect against the 3-NPA-induced decline in ovarian function by activating the SESTRIN2–NRF2 pathway. In ovarian oxidative injury, activation of the SESTRIN2–NRF2 pathway reduces ROS accumulation, supports mitochondrial protection, and helps preserve ovarian function (Huang et al., 2024). These findings link SESN2 to natural antioxidant-mediated ovarian protection.
Figure 4. Sestrin2–NRF2 signaling in ovarian oxidative-stress protection.
(Huang et al., 2024)
1. Methods for Detecting and Analyzing SESN2
SESN2 can be evaluated at the mRNA, protein, and functional levels. RT-qPCR is most commonly used to evaluate SESN2 gene expression, while Western blotting assesses SESN2 protein abundance (Huang et al., 2024). Immunohistochemistry and immunofluorescence are typically used to localize SESN2 to specific tissues or cellular compartments. Functional studies often require SESN2 knockdown or overexpression. These approaches help determine whether SESN2 is only associated with disease or whether it directly contributes to protection or injury (Xi et al., 2024). Mechanistic studies commonly combine SESN2 knockdown or overexpression with ROS assays, apoptosis analysis, mitochondrial membrane-potential detection, autophagy markers, and ferroptosis-related readouts. In addition to assessing antioxidant activity, endpoints for reproductive research studies involving SESN2 may include granulosa cell viability, mitochondrial function, apoptosis, ovarian histology, hormone levels, and oxidative stress biomarkers (Wang et al., 2023). These endpoints can help assess whether SESN2 improves ovarian-cell survival and protects against ovarian functional decline. In natural-compound studies, SESN2 can also be tested as a pathway target for antioxidants such as proanthocyanidins (Huang et al., 2024).
2. Therapeutic Potential and Future Perspectives
SESN2’s therapeutic effects arise from its position upstream of multiple protective mechanisms. Activation of SESN2 may mitigate oxidative stress, ferroptosis, inflammation, mitochondrial damage, and endoplasmic reticulum (ER) stress (Lu et al., 2023). Thus, SESN2 is better described as a potential therapeutic target that still requires disease-specific validation (Rooban et al., 2024; Wang et al., 2024). In addition to metabolic disease, SESN2 may be involved in regulating insulin sensitivity, lipid metabolism, and energy balance. Its ability to influence AMPK, mTORC1, and autophagy provides relevance to diabetes, obesity, fatty liver disease, and cardiac injury (Gong et al., 2021). However, because SESN2 has varying effects depending on the specific tissue or disease condition, careful consideration is required when designing therapeutic approaches. In reproductive biology, SESN2 should be investigated further using models that connect granulosa-cell survival, oocyte competence, ovarian reserve, and fertility outcomes under oxidative stress (Wang et al., 2024; Wang et al., 2023; Yang et al., 2024). It would also be valuable to test whether natural compounds can protect ovarian function through SESN2–NRF2, SESN2–AMPK, or SESN2–mTOR pathways (Huang et al., 2024).
3. Conclusion
SESN2 is an important stress-responsive protein that helps maintain cellular homeostasis during oxidative, metabolic, and organelle-related injury. It supports antioxidant defense and regulates AMPK-mTOR signaling, autophagy, mitophagy, ferroptosis, and endoplasmic reticulum (ER) quality control. Through these mechanisms, SESN2 can reduce ROS production, limit lipid peroxidation, promote the removal of damaged organelles, and improve cellular adaptation to stress. Its reproductive relevance is also important because ovarian function depends on mitochondrial activity, granulosa cell survival, and redox stability. SESN2-regulated pathways may therefore contribute to ovarian protection under oxidative stress, although direct effects on fertility outcomes still require further confirmation.
References
Abdelsalam, S.S., Zahid, M.A., Raïq, H., Abunada, H., Elsayed, A.E., Parray, A. and Agouni, A. (2024). The association between plasma levels of Sestrin2 and risk factors of cardiovascular diseases in healthy and diabetic adults: a study of Qatar Biobank data. Biomolecules and Biomedicine, 25(7), pp.1479–1490. https://doi.org/10.17305/bb.2024.11418.
Chen, F., Kang, R., Tang, D. and Liu, J. (2024). Ferroptosis: principles and significance in health and disease. Journal of Hematology & Oncology, 17, 41. https://doi.org/10.1186/s13045-024-01564-3.
De Leonibus, C., Maddaluno, M., Ferriero, R., Besio, R., Cinque, L., Lim, P.J., Palma, A., De Cegli, R., Gagliotta, S., Montefusco, S., Iavazzo, M., Rohrbach, M., Giunta, C., Polishchuk, E., Medina, D.L., Di Bernardo, D., Forlino, A., Piccolo, P. and Settembre, C. (2024). Sestrin2 drives ER-phagy in response to protein misfolding. Developmental Cell, 59, pp.2035–2052.e10. https://doi.org/10.1016/j.devcel.2024.07.004.
Dou, X., Dong, W., Gu, Y., Zhang, T. and Zhang, J. (2023). Significance of serum sestrin2 as a biomarker of severity and functional outcome in acute intracerebral hemorrhage: a prospective observational longitudinal study. BMC Neurology, 23, 424. https://doi.org/10.1186/s12883-023-03470-6.
Gong, L., Wang, Z.Z., Wang, Z.G. and Zhang, Z. (2021). Sestrin2 as a potential target for regulating metabolic-related diseases. Frontiers in Endocrinology, 12, 751020. https://doi.org/10.3389/fendo.2021.751020.
Haidurov, A. and Budanov, A.V. (2024). Locked in structure: Sestrin and GATOR—a billion-year marriage. Cells, 13, 1587. https://doi.org/10.3390/cells13181587.
Huang, Y., Cui, Y., Huang, J., Huang, X., Wang, Z., Luo, T. and Li, J. (2024). Proanthocyanidins protects 3-NPA-induced ovarian function decline by activating SESTRIN2-NRF2-mediated oxidative stress in mice. Scientific Reports, 14, 25643. https://doi.org/10.1038/s41598-024-76743-w.
Ju, W., Zhao, Y., Yu, Y., Zhao, S. and Lian, F. (2024). Mechanisms of mitochondrial dysfunction in ovarian aging and potential interventions. Frontiers in Endocrinology, 15, 1361289. https://doi.org/10.3389/fendo.2024.1361289.
Li, B., Ming, H., Qin, S., Nice, E.C., Dong, J., Du, Z. and Huang, C. (2025). Redox regulation: mechanisms, biology and therapeutic targets in diseases. Signal Transduction and Targeted Therapy, 10, 72. https://doi.org/10.1038/s41392-024-02095-6.
Liu, X., Tuerxun, H., Zhao, Y.X., Li, Y., Wen, S., Li, X. and Zhao, Y.G. (2025). Crosstalk between ferroptosis and autophagy: broaden horizons of cancer therapy. Journal of Translational Medicine, 23, 18. https://doi.org/10.1186/s12967-024-06059-w.
Lu, C., Jiang, Y., Xu, W. and Bao, X. (2023). Sestrin2: multifaceted functions, molecular basis, and its implications in liver diseases. Cell Death & Disease, 14, 160. https://doi.org/10.1038/s41419-023-05669-4.
Ro, S.-H., Fay, J., Cyuzuzo, C.I., Jang, Y., Lee, N., Song, H.-S. and Harris, E.N. (2020). SESTRINs: emerging dynamic stress-sensors in metabolic and environmental health. Frontiers in Cell and Developmental Biology, 8, 603421. https://doi.org/10.3389/fcell.2020.603421.
Rooban, S., Senghor, K.A.A., Vinodhini, V.M. and Kumar, J.S. (2024). Sestrin2 at the crossroads of cardiovascular disease and diabetes: a comprehensive review. Obesity Medicine, 51, 100558. https://doi.org/10.1016/j.obmed.2024.100558.
Valenstein, M.L., Wranik, M., Lalgudi, P.V., Linde-Garelli, K.Y., Choi, Y., Chivukula, R.R., Sabatini, D.M. and Rogala, K.B. (2025). Structural basis for the dynamic regulation of mTORC1 by amino acids. Nature, 646, pp.493–500. https://doi.org/10.1038/s41586-025-09428-7.
Wang, M., Chen, W., Zeng, X., Wang, T., Sun, Y. and Yang, Q. (2024). Sestrin1, 2, and 3 are dispensable for female fertility in mice. Journal of Ovarian Research, 17, 28. https://doi.org/10.1186/s13048-024-01345-z.
Wang, X., Yang, J., Li, H.Y., Mu, H., Zeng, L., Cai, S., Su, P., Li, H.B., Zhang, L. and Xiang, W. (2023). miR-484 mediates oxidative stress-induced ovarian dysfunction and promotes granulosa cell apoptosis via SESN2 downregulation. Redox Biology, 62, 102684. https://doi.org/10.1016/j.redox.2023.102684.
Wu, S.A., Li, Z.J. and Qi, L. (2025). Endoplasmic reticulum (ER) protein degradation by ER-associated degradation and ER-phagy. Trends in Cell Biology, 35, pp.576–591. https://doi.org/10.1016/j.tcb.2025.01.002.
Xi, X., Chen, Q., Ma, J., Wang, X., Zhang, J. and Li, Y. (2024). Sestrin2 ameliorates diabetic retinopathy by regulating autophagy and ferroptosis. Journal of Molecular Histology, 55, pp.169–184. https://doi.org/10.1007/s10735-023-10180-3.
Yang, M., Wei, X., Yi, X. and Jiang, D.-S. (2024). Mitophagy-related regulated cell death: molecular mechanisms and disease implications. Cell Death & Disease, 15, 505. https://doi.org/10.1038/s41419-024-06804-5.
Yang, Z., Liu, S. and Pan, X. (2024). Research progress on mitochondrial damage and repairing in oocytes: a review. Mitochondrion, 75, 101845. https://doi.org/10.1016/j.mito.2024.101845.
Zhang, L., Ding, K., Liao, S., Zhang, Y., Liao, H., Chen, R. and Meng, Q. (2024). Sestrin2 reduces ferroptosis via the Keap1/Nrf2 signaling pathway after intestinal ischemia-reperfusion. Free Radical Biology and Medicine, 214, pp.115–128. https://doi.org/10.1016/j.freeradbiomed.2024.02.003.








